Structure and Function of Amino Acids was adapted from Kohlmeier, M. (2015). Nutrient metabolism: Structures, functions, and genes (2nd Edition). Academic Press. Manual annotation have been made and citations and references are duly provided.
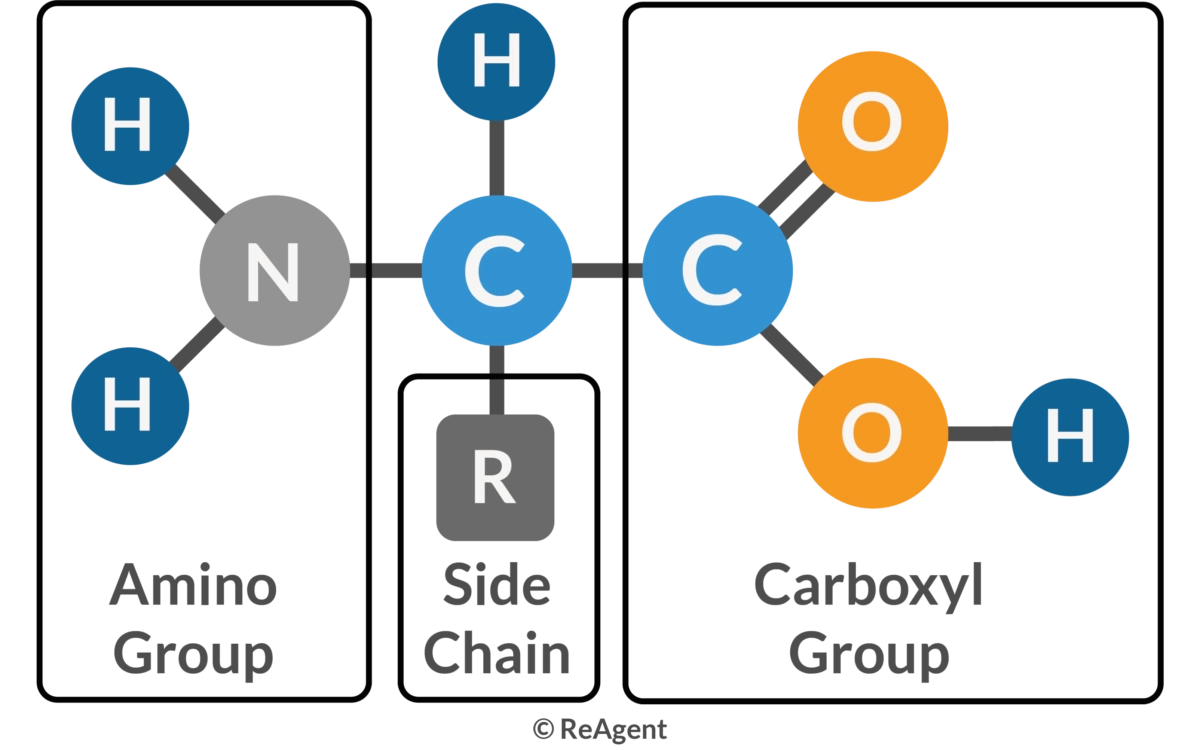
Amino acids are organic molecules that serve as the building blocks of proteins. Each amino acid consists of a central α-carbon atom bonded to a basic amino group (−NH2), an acidic carboxyl group (−COOH), a hydrogen atom, and a unique organic R group (side chain). Proteins, which are essential for catalyzing cellular chemical reactions, providing structural elements, and binding cells into tissues, are composed of these amino acids. In humans, there are 20 standard amino acids. Nine of these are essential and must be obtained through the diet: histidine, isoleucine, leucine, lysine, methionine, phenylalanine, threonine, tryptophan, and valine. Five amino acids are nonessential, as the body can synthesize them: alanine, asparagine, aspartic acid, glutamic acid, and serine. The remaining six are conditional amino acids, essential only during certain life stages or under specific conditions: arginine, cysteine, glutamine, glycine, proline, and tyrosine. Additionally, selenocysteine, sometimes regarded as the 21st amino acid, is derived from serine during protein biosynthesis.
Amino acids are the building blocks of protein synthesis. They are structural elements and energy sources of cells necessary for normal cell growth, differentiation and function. Amino acid metabolism disorders have been linked with a number of pathological conditions, including metabolic diseases, cardiovascular diseases, immune diseases, and cancer.
The amino acids relevant for human nutrition comprise dozens of compounds with a broad variation of characteristics and functions. Amino acids, in the form of proteins of connective tissues and muscles, hold the body together and give it strength to move around. Many other proteins catalyze reactions, transport compounds, and coordinate many events. Amino acids and their derivatives also serve virtually all categories of signaling, metabolism, and functional support.
COMPOSITION AND STRUCTURE
By definition, amino acids are always made up of an amino group, an acid, and a side chain. The amino group is most often adjacent to the primary carboxyl group (alpha-position), but may be in the betaposition instead (e.g., in beta-alanine). As in carnitine, a tertiary amine may serve the same function. Similarly, the acid group may be represented by a sulfoxy function, as in taurine. The side chain may be just a single hydrogen (in glycine), straight (alanine), or branched (valine, leucine, and isoleucine) aliphatic chains, contain aromatic rings (phenylalanine, tyrosine, and tryptophan), sulfur (methionine, cysteine, and taurine), selenium (selenocysteine), hydroxyl groups (serine, threonine, hydroxyproline, and hydroxylysine), second carboxyl groups (glutamate and aspartate) or third carboxyl groups (gamma-carboxyl glutamate and gammacarboxyl aspartate), amido groups (glutamine and asparagine), or additional nitrogen-containing groups (lysine, arginine, and histidine).
Basic Structure of amino acids illustrating: A carbon (the alpha carbon), hydrogen atom (H), Carboxyl group (-COOH), Amino group (-NH2) and "R / Side chain" group which mainly determines amino acid's physicla and chemical properties
Classification
Amino acids
Essential amino acids cannot be synthesized by humans, require dietary intake; Non-essential are synthesed by humans using various metabolic pathways; Semi-essential amino acids, also known as conditionally essential amino acids, are typically synthesize under normal circumstances, but under certain conditions, the body may require additional dietary intake.
Since the alpha-carbon is chiral for all amino acids except glycine, two stereoisomeric conformations are possible. When the carboxylic group in structural diagrams is depicted at the top and the side chain at the bottom, the l form is the one that has the amino group pointing to the left, the d form has it pointing to the right. Human proteins contain only l-amino amino acids (and glycine). d-Amino acids are quantitatively less common in mammals than in fungi and bacteria, but a few (including d-aspartate and d-serine) are synthesized by humans (D’Aniello et al., 1993; Wolosker et al., 1999, 2000).
Some amino acids form only a few specific peptides, such as taurine as part of glutaurine, and betaalanine as part of carnosine and anserine. Several other amino acids are not a regular part of human proteins but serve specific important functions. This is the case with carnitine (fatty acid transport), taurine (osmolyte, neuronal agent, and antioxidant), ornithine (urea cycle and polyamine synthesis), and citrulline (urea cycle)
Name and Formula
Side Chain
2D Structure
ENDOGENOUS SOURCES
Precursors from intermediary metabolism: Several amino acids are synthesized through the addition of an amino group to a common alpha-keto acid. Alpha-ketoglutarate and oxaloacetate are readily available Krebs-cycle intermediates, and pyruvate is a glycolysis product. Glyoxylate is a much less abundant intermediary metabolite, which arises from the metabolism of hydroxyproline, ethanolamine, glycolate, ethylene glycol, and a few other minor sources. A slightly more complex case is serine, which is used to synthesize 3-phosphoglycerate in a three-step process. The main source of the amino groups is other amino acids. The exchange between glutamate and aspartate (aspartate aminotransferase, EC2.6.1.1) and glutamate and alanine (alanine aminotransferase, EC2.6.1.2) is particularly important, as underscored by the abundance of the respective aminotransferases in the muscle, liver, and other tissues. The branched-chain amino acids (BCAAs) leucine, valine, and isoleucine are another abundant source of amino groups for transamination in muscle, especially in the postprandial phase. All of these aminotransferases require pyridoxal 5-phosphate (PHP) as a covalently bound cofactor. Ammonium ions can also contribute directly to amino acid synthesis, but the extent is limited. Glutamate dehydrogenase (EC1.4.1.3) catalyzes the nicotine adenine dinucleotide phosphate (NADPH)–dependent amidation of alpha-ketoglutarate in mitochondria. Alternatively, ammonia can also be linked to glutamate by glutamate dehydrogenase (EC1.4.1.3) in a distinct, NADPH-dependent reaction. Ammonia in cytosol, on the other hand, is used by glutamate-ammonia ligase (EC6.3.1.2) for glutamate amidation in an adenosine triphosphate (ATP)–energized reaction.
Amino acids as precursors: Tyrosine, cysteine, glutamine, asparagine, arginine, ornithine, citrulline, proline, taurine, and carnitine are derived from other amino acids, some of which are essential and must come from food. Tyrosine can be derived from the essential amino acid phenylalanine by hydroxylation. Cysteine is synthesized via the trans-sulfuration pathway from methionine. Glutamine and asparagine are the amidation products of their respective dicarboxylate amino acid precursors. Arginine, ornithine, citrulline, and proline start from glutamate; net synthesis occurs only in the intestinal wall. The precursor for routine synthesis is cysteine
Posttranslational modification: The precursors of a few amino acids are incorporated into specific proteins and modifications take place only during or after protein synthesis. Many amino acid residues in proteins are modified, and they often acquire full functionality only after such changes. Common modifications include phosphorylation, acylation, and glycosylation. However, after breakdown of a protein, most of the modified amino acids can still be reutilized for protein synthesis. This is not true for a small group of amino acids that are permanently altered by modification of the protein. This group includes hydroxyproline, hydroxylysine, methylhistidine, gamma-carboxyglutamate, gamma-carboxyaspartate, and carnitine. The posttranslational hydroxylation of several procollagens and elastins generates hydroxyproline and hydroxylysine. Methylhistidine arises from the posttranslational methylation of actin and myosin in muscle and the synthesis of anserine and a few other specific peptides. A vitamin K–dependent reaction carboxylates the γ-carbon of specific glutamate residues in several coagulation factors and a few other proteins. Analogous reactions with aspartate residues may also occur. The selenocysteine residues, which are constituents of the reactive centers in a few enzymes, are produced by a complex series of reactions from the serine precursor. Serine-tRNA ligase (EC6.1.1.11) charges tRNA(Ser)sec with serine. The pyridoxal phosphate-dependent l-seryl-tRNAsec selenium transferase (EC2.9.1.l) then substitutes the hydroxyl group of the serine with selenophosphate. The tRNA is now ready to add its load of selenocysteine to an emerging peptide strand in an unusually complex process. Carnitine synthesis is similarly complicated. In this case, three methyl groups are added to specific lysines in myosin, actin, histones, and other proteins by histone–lysine N-methyltransferase (EC2.1.1.43). After the proteins have been broken down in the course of their normal turnover, carnitine synthesis requires another four steps for completion
DIETARY SOURCES
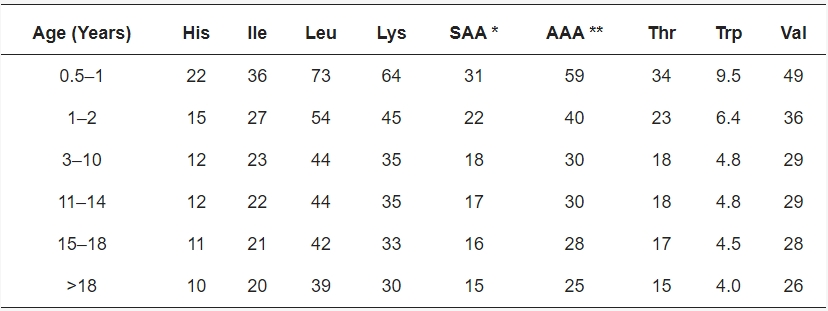
Most amino acids are consumed with a wide variety of proteins and peptides in foods of animal and plant origin. These foods provide all 20 amino acids used for making the body’s own proteins, peptides, and other compounds that require amino acids as precursors. Phenylalanine, tryptophan, methionine, histidine, lysine, valine, leucine, isoleucine, and threonine are called essential because they must be provided with food. Current recommendations are to consume daily at least 0.8g of protein per kilogram body weight, and more during pregnancy (Food and Nutrition Board, Institute of Medicine, 2005). Children (Elango et al., 2011) and some adults (Kohlmeier, 2012) may have significantly higher requirements than suggested by current recommendations
Recommended intake of indispensable amino acids (in mg/kg body weight/day) for humans in different age groups. * SAA = Sulphur-containing amino acids (Cys + Met); ** AAA = Aromatic amino acids (Phe + Tyr). (Adhikari et al., 2022)
Several other amino acids, which are not incorporated in human proteins, have nutritional value nonetheless as nonessential precursors for the endogenous synthesis of important compounds, or just as a source of energy; examples include carnitine, taurine, and ornithine. The unusual amino acid l-theanine (5-N-ethylglutmine) constitutes half of the amino acids in tea (Camellia sinensis) leaves and appears to confer some beneficial effects (Unno et al., 2013). Finally, there are amino acids that cannot be utilized well or even are harmful, such as some d-amino acids. Particular peptides, of course, may have their own specific effects. Some are potent hormones (though most will not be taken up intact and active); others may be potent toxins (e.g., amanitin from death cap mushrooms).
Cooking of foods tends to improve the overall bioavailability of protein and confers desirable flavors and aromas. However, heating can modify amino acids in numerous ways and may lead to significant losses of cysteine, methionine, threonine, serine, and tryptophan (Dworschak, 1980). Heating also can promote the cross-linking of l-alanine with other amino acids in food proteins (generating lysinoalanine, ornithinoalanine, histidinoalanine, and phenylethylaminoalanine), the formation of dehydroalanine, methyldehydroalanine, beta-aminoalanine, and racemization to d-amino acids (Friedman, 1999). Another typical effect of heating is the cross-linking of amino acids and sugars. The attractive browning that develops during the cooking, frying, or baking of foods reflects these reactions. Initially, Schiff bases form between lysines, arginines, and sugars, which rearrange to more stable Amadori products (Biemel et al., 2001). With continued heating, these intermediates react further and form insoluble protein complexes with extensive lysine-arginine cross-links. These complexes are commonly referred to as Maillard products or advanced glycation end products. Presumably due to their low solubility and the extensive chemical modifications of their amino acid constituents, such Maillard products have extremely low bioavailability and food value (Erbersdobler and Faist, 2001).
Another problem related to heating foods to high temperatures, such as with broiling, is the formation of cancer-inducing compounds. Condensation and pyrolysis of creatine with aromatic amino acids (2-amino-1-methyl-6-phenylimidazo[4,5-b] pyridine, PhIP), glycine (2-amino-3,8-dimethylimidazo[4,5- f]quinoxaline, MeIQx), or other amino acids generate very potent carcinogens (Oguri et al., 1998; Schut and Snyderwine, 1999). These compounds readily attach to DNA (adduct formation) and cause mutations
Daily protein intake in the United States declines with age, from about 68g in young women to 60g in women over the age of 60 (Smit et al., 1999). Similar trends are seen for men, though at overall higher intake levels.
Foods that contain all nine essential amino acids are referred to as complete proteins. The following foods are complete protein sources: meat, seafood, poultry, eggs, dairy products. Soy and pea protein are plant-based complete protein sources. Other plant-based sources of protein, such as beans, nuts, and certain grains, are considered incomplete proteins because they lack one or more of the essential amino acids. However, if you’re following a plant-based diet, you can still ensure proper intake of all nine essential amino acids by eating a variety of plant proteins each day. For example, choosing a variety of plant-based proteins, such as beans, nuts, seeds, whole grains, and vegetables, can ensure that you meet your essential amino acid needs, even if you choose to exclude animal products from your diet.
The nutritional value or quality of structurally different proteins varies and is governed by amino acid composition, ratios of essential amino acids, susceptibility to hydrolysis during digestion, source, and the effects of processing. To optimize the biological utilization of proteins, a better understanding is needed of the various interrelated parameters that influence their nutritive value. This review attempts to contribute to this effort. ...... Recent studies on the nutritional quality of more than 50 common and uncommon protein sources including cereals, legumes, other seeds, meat, seafood, insects, leaves, mushrooms, and potatoes are reviewed. Also described are advantages of consuming low-quality proteins fortified with essential amino acids, nutritional benefits of mixtures of complementary protein sources, plant genetic approaches to improving the nutritive value of foods, problems associated with liquid diets for adults and infants, socioeconomic aspects of new protein foods, and the influence of protein type and quality on lactation, the immune system, and serum lipids ....."
DIGESTION
The chewing and mixing with saliva in the mouth (mastication) breaks solid food into small particles and initiate digestion. The importance of oral protein-cleaving enzymes appears to be minimal, however. Predigestion in the stomach initiates protein digestion. The acidity of the added hydrogen chloride denatures ingested proteins. Pepsin (EC3.4.23.1) and gastricsin (pepsinogen C, EC3.4.23.3) hydrolyze ingested proteins with broad specificity.
A second tier of protein-digesting enzymes comes from the pancreas. These include several forms of trypsin, alpha-chymotrypsin, chymotrypsin C, carboxypeptidase B, and two forms each of elastase II, endopeptidase E (EC3.4.21.70), and carboxypeptidase A (EC3.4.2.1). Like most pancreas enzymes, these proteases are not active when they reach the small intestine. Duodenal glands secrete duodenase (EC3.4.21. B3, Zamolodchikova et al., 2000). This serine protease activates the brush border protease enteropeptidase (enterokinase, EC3.4.21.9) by cleaving it. Enterokinase, in turn, cleaves and activates trypsin (EC3.4.21.4), and trypsin finally activates the other pancreas proteases. The activity of the protease at the top of this activating cascade is inhibited by alpha 1-proteinase inhibitor (Gladysheva et al., 2001). Additional proteases may have more narrow specificity. One such enzyme is tissue kallikrein (EC3.4.21.35), a kinin activator, which is expressed in acinar cells together with its inhibitor kallistatin (Wolf et al., 1998)
The third tier of enzymes works near or directly at the brush border membrane on a mixture that by now contains relatively small peptides. This means that individual amino acids, dipeptides, or tripeptides are released predominantly near the surface of the brush border membrane, from where they can be taken up into the epithelial cell layer. A few aminopeptidases, particularly membrane alanine aminopeptidase (aminopeptidase N, EC3.4.11.2), are relatively abundant. This enzyme releases N-terminal alanine and a broad spectrum of other amino acids from peptides, amides, or arylamides. There is a considerable diversity of enzymes commensurate with the different substrates. An example of an enzyme with a relatively narrow mission is folylpoly-γ-glutamate carboxypeptidase, which converts food folate into the absorbable free form. The many people who have a slightly less active form absorb food folate less well and are more likely to suffer detrimental consequences of low intake (Devlin et al., 2000).
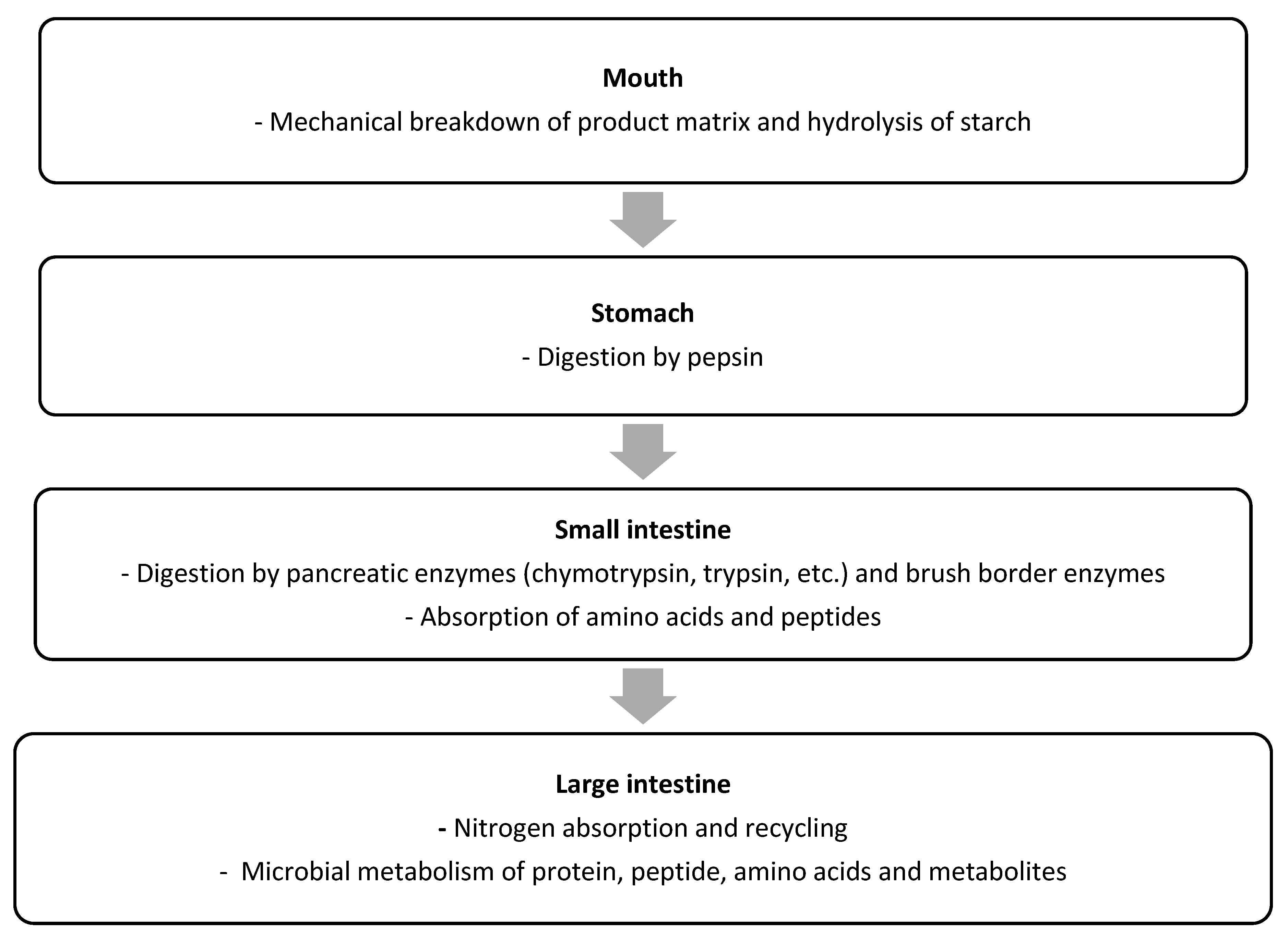
Protein Digestion and Absorption by Humans
Proteins are the key source of indispensable amino acids (IAAs) for the human body. However, for the IAAs, as well as the DAAs, from a protein to be utilized by the human body, the protein first needs to be hydrolysed into free amino acids and small (di- and tri-) peptides, which can subsequently be absorbed in the bloodstream. Protein digestion is a complex, multistage process, as schematically outlined in Figure. The protein digestion process begins with the mechanical disruption of the product matrix containing the protein during mastication in the mouth. While protein breakdown does not occur at this stage, the oral phase of digestion can be important for protein digestion, because by disrupting the product matrix, the surface area increases, which increases exposure of the protein to digestive proteases and peptidases during the later stages of digestion. Digestion of starch by salivary amylase can also disrupt the product structure, thereby increasing accessibility of protein to digestive enzymes in later stages of the digestion process.
Schematic overview of the key steps of protein digestion and absorption in humans. (Adhikari et al., 2022)
Following the oral phase of digestion, which typically lasts only for a short time (less than a minute), the product passes on to the stomach, where the gastric phase of the digestion process takes place. In this phase, the product is mixed with gastric juice, which has a low pH (1–2 for adults) and contains the protease pepsin. The mixing of gastric juice and product is facilitated through contractions of the stomach. In the stomach, some hydrolysis of proteins by pepsin occurs, resulting in (poly)peptide formation, but complete protein digestion to free amino acids and peptides small enough for absorption does not occur at this stage
Following gastric digestion, the chyme is delivered to the duodenum at a rate determined by the gastric emptying. In the small intestine, the chyme is mixed with pancreatic proteases and peptidases, such as trypsin, chymotrypsin, and carboxypeptidase A. Together with intestinal brush border enzymes, these enzymes hydrolyse the proteins and (poly)peptides into amino acids, di-, tri-, and oligopeptides. Pancreatic proteases and peptidases are considered rigorous compared to pepsin, and most of the protein digestion occurs in the small intestine rather than in the stomach [46]. The amino acids and di- and tripeptides that are released can be taken up across the small intestinal mucosa and are generally considered to be almost fully absorbed by the end of small intestine, i.e., the terminal ileum. The amino acids and peptides not absorbed at the terminal ileum pass to the large intestine. The large intestine, especially the caecum, also contains amino acid transporters, but there is thus far no evidence that the absorption of amino acids in large intestine occurs in relevant quantities. If fully absorbed, the amino acids absorbed in the large intestine in pigs would increase the level of total amino acids absorbed by only 0.1% for whey protein and by 3.5% for zein. The quantity of amino acids passing to large intestine is thus only affected by the absorption of amino acids and small peptides from the consumed protein to a limited amount. The proportion of unabsorbed amino acids and peptides, as well as of undigested protein and polypeptides, can also be digested and fermented by the microbiota. Furthermore, colonocytes are capable of synthesizing and metabolizing amino acids which are likely derived through blood circulation rather than the digesta. Given that protein is mainly digested and absorbed in the small intestine, and microbial protein is formed in the large intestine, digesta samples collected from either site may differ notably, and cause variation in measurements of protein digestibility ....
Proteolytic enzymes (proteases) degrade dietary proteins into their constituent amino acids in the stomach and intestine. Digestive proteases are synthesized as larger, inactive forms (zymogens), which, after secretion, are cleaved to produce active proteases. The amino acids indicated after the curly brace in the diagram below are the preferred amino acids at which each of the indicated enzymes cleaves.
In the stomach, pepsin begins the digestion of dietary proteins by hydrolyzing them to smaller polypeptides. Pepsinogen is secreted by chief cells of the stomach, parietal cells secrete HCl. The acid environment alters the conformation of pepsinogen so that it can cleave itself to yield pepsin. Pepsin acts as an endopeptidase to cleave dietary proteins with a broad spectrum of specificity, although it prefers to cleave peptide bonds in which the carboxyl group is provided by aromatic or acidic amino acids. The products are smaller peptides and some free amino acids
In the intestine, bicarbonate neutralizes stomach acid, and the pancreas secretes several inactive proenzymes (zymogens), which, when activated, collectively digest peptides to single amino acids. Enteropeptidase, secreted by the brush border cells of the small intestine cleaves trypsinogen to yield the active serine protease trypsin. Trypsin cleaves inactive chymotrypsinogen to yield active chymotrypsin, inactive proelastase, to yield active elastase, and inactive procarboxypeptidases to yield active carboxypeptidases. Thus, trypsin plays a central role because it cleaves dietary proteins and activates other proteases that cleave dietary protein. Each protease exhibits cleavage specificity: trypsin cleaves at the carboxy side of arg and lys; chymotrypsin cleaves at the carboxy side of phe, tyr, trp and leu; elastase cleaves at the carboxy side of ala, gly and ser. Carboxypeptidase A cleaves single amino acids from the carboxyl terminus, with a specificity for hydrophobic and branched side chain amino acids; carboxypeptidase B cleaves single amino acids from the carboxyl terminus, with a specificity for basic (arg and lys) amino acids. Aminopeptidases, located on the brush border, cleave one amino acid at a time from the amino end of peptides. Intracellular peptidases cleave small peptides absorbed by cells.
ADSORPTION
After ingestion, proteins are denatured by the acid in the stomach, where they are also cleaved into smaller peptides by the enzyme pepsin, which is activated by the increase in stomach acidity that occurs on feeding. The proteins and peptides then pass into the small intestine, where the peptide bonds are hydrolyzed by a variety of enzymes. These bond-specific enzymes originate in the pancreas and include trypsin, chymotrypsins, elastase, and carboxypeptidases. The resultant mixture of free amino acids and small peptides is then transported into the mucosal cells by a number of carrier systems for specific amino acids and for di- and tri-peptides, each specific for a limited range of peptide substrates. After intracellular hydrolysis of the absorbed peptides, the free amino acids are then secreted into the portal blood by other specific carrier systems in the mucosal cell or are further metabolized within the cell itself. Absorbed amino acids pass into the liver, where a portion of the amino acids are taken up and used; the remainder pass through into the systemic circulation and are utilized by the peripheral tissues. /Amino acids/
DISTRIBUTION
We're working on it, check update later
METABOLISM
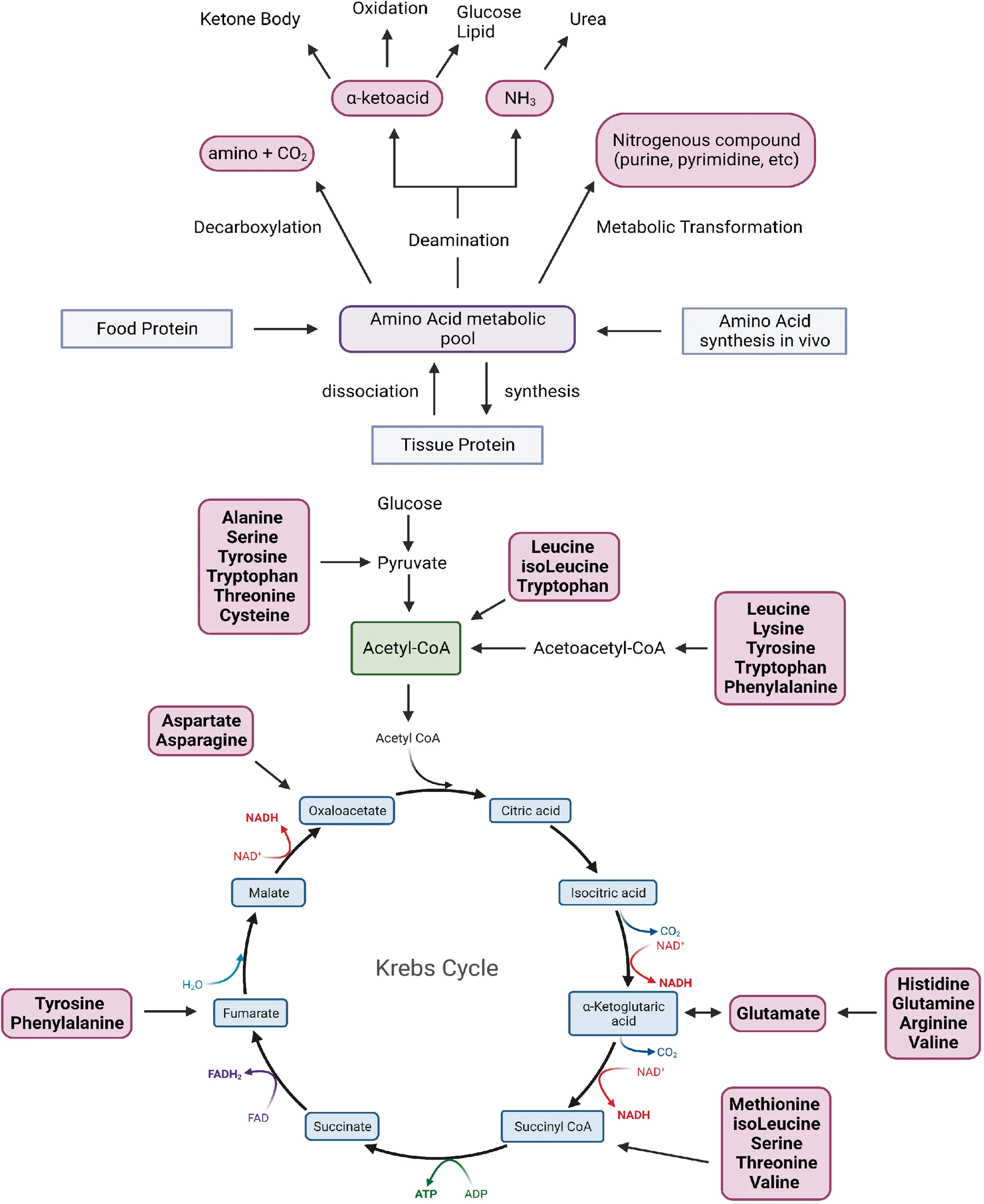
One of the first steps in the breakdown of amino acids is the removal of the amino group. Typically, this involves a transamination reaction, often with alpha-ketoglutarate as the acceptor. All transaminases require PHP as a covalently bound cofactor. Ammonia may also be released directly, such as in the deamination reaction catalyzed by d-amino acid oxidase (EC1.4.3.3), which occurs especially in the kidneys (Hasegawa et al., 2011). The breakdown of a few amino acids involves nothing more than the reversal of the reactions responsible for their synthesis. Glutamate, for example, may shed its amino group in any of numerous possible transamination reactions, and the resulting alpha-ketoglutarate can be utilized through the Krebs cycle. Some other amino acids require a much larger number of reactions and may depend on several vitamin cofactors. The complete oxidation of tryptophan, for example, takes more than 20 steps and requires adequate supplies of thiamin, riboflavin, vitamin B6, niacin, pantothenate, lipoate, ubiquinone, iron, and magnesium.
Most of the amino acids in proteins (alanine, valine, isoleucine, proline, phenylalanine, tyrosine, methionine, cysteine, serine, threonine, aspartate, glutamate, glutamine, aspartate, asparagine, histidine, and arginine) can be converted into glucose and are referred to as glucogenic; therefore, isoleucine, lysine, phenylalanine, tyrosine, and tryptophan are considered ketogenic because their catabolism generates ketone bodies or their precursors (acetoacetate, acetate, or acetyl-CoA)
Overview of amino acid metabolism.
Glycine: Most glycine is converted in the mitochondria into serine (using 5,10-methylene tetrahydrofolate) or undergoes deamination, decarboxylation, and one-carbon transfer to folate (generating 5,10-methylene tetrahydrofolate). Much smaller amounts are converted into glyoxylate. Glycine is used for purine nucleotide synthesis. Glycine breakdown requires riboflavin, vitamin B6, niacin, folate, lipoate, ubiquinone, iron, and magnesium.
l-Alanine: Transamination generates pyruvate, which can be metabolized to acetyl-CoA (which would make alanine ketogenic), or used for glucose synthesis. d-alanine can also be utilized after conversion to pyruvate (Ogawa and Fujioka, 1981) by glycine hydroxymethyltransferase (EC2.1.2.1). Complete alanine breakdown requires thiamin, riboflavin, vitamin B6, niacin, lipoate, ubiquinone, iron, and magnesium.
l-Proline: The bulk of proline is broken down via glutamate to alpha-ketoglutarate, which can then be used for glucose synthesis or further metabolism through the Krebs cycle. A very significant amount is used in the intestines as a precursor for the synthesis of citrulline, ornithine, and arginine. The oxidation of proline is remarkable for its generation of oxygen free radicals (Donald et al., 2001). Complete oxidation of proline depends on thiamin, riboflavin, vitamin B6, niacin, lipoate, ubiquinone, iron, and magnesium. Only the kidneys metabolize significant amounts of hydroxyproline.
l-Valine: A total of 10 reactions is required to metabolize valine to succinyl-CoA, which can then be utilized further through the Krebs cycle and oxidative phosphorylation. Complete oxidation requires thiamin, riboflavin, niacin, vitamin B6, vitamin B12, pantothenate, biotin, lipoate, ubiquinone, iron, and magnesium.
l-Leucine: Most leucine is metabolized to acetoacetate and acetyl-CoA. This makes leucine the main ketogenic amino acid. About 5–10% is oxidized via beta-hydroxy beta-methylbutyrate (HMB) to acetyl-CoA. Even smaller amounts (mainly in the testis) are converted in an adenosylcobalamindependent reaction to beta-leucine. Normal leucine metabolism uses thiamin, riboflavin, vitamin B6, niacin, vitamin B12, biotin, pantothenate, lipoate, ubiquinone, iron, and magnesium.
l-Isoleucine: This BCAA is broken down to succinyl-CoA and acetyl-CoA in six steps. Utilization depends on adequate availability of thiamin, riboflavin, niacin, vitamin B6, vitamin B12, pantothenate, biotin, lipoate, ubiquinone, iron, and magnesium.
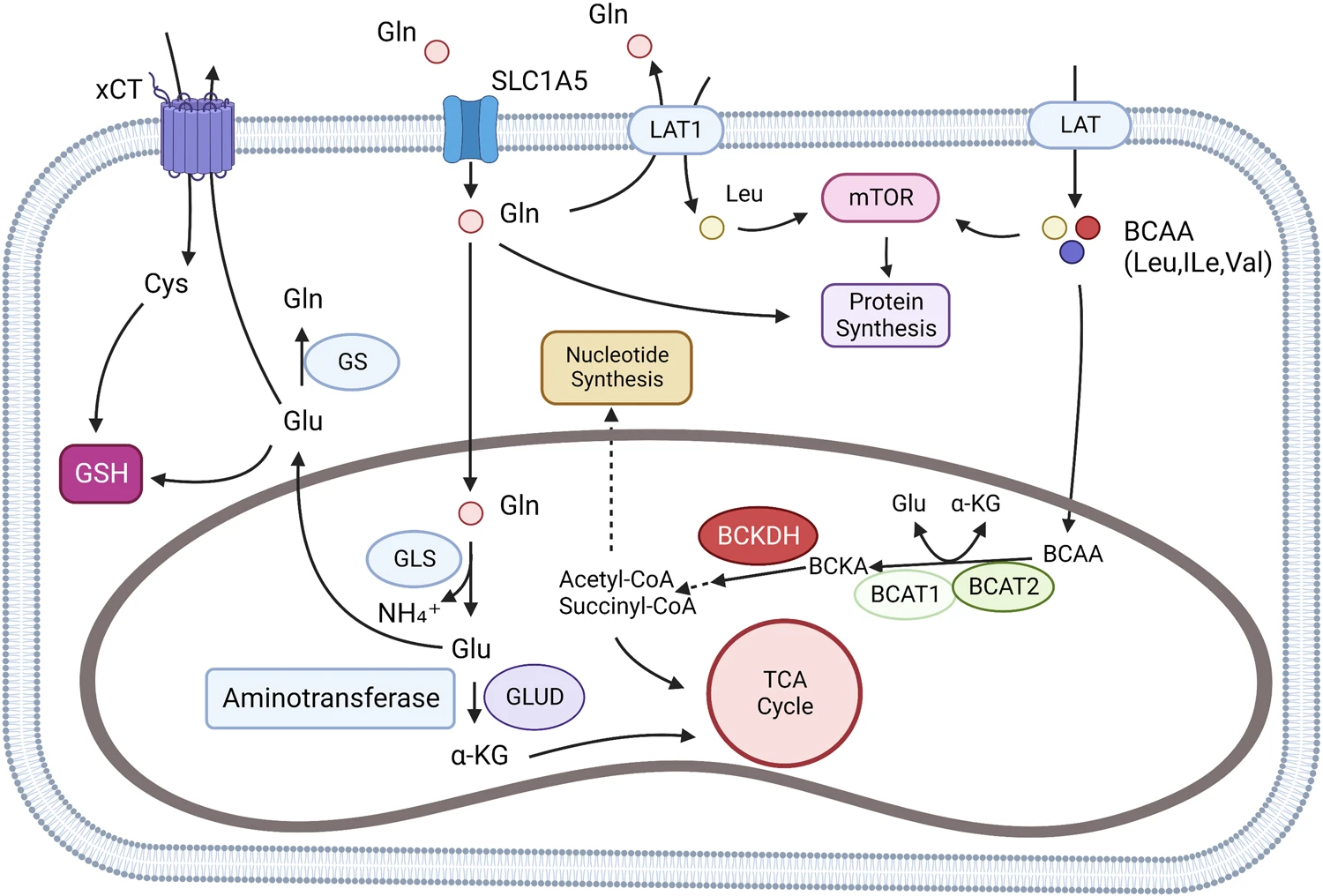
Glutamine and BCAA metabolism. BCAAs can be absorbed by the cell through L-type amino acid transporter (LATs), and L-type amino acid transporter 1(LAT1) can also exchange intracellular glutamine with extracellular leucine. In cells, BCAAs are catalyzed to formα-ketoisocaproate (KIC), α-ketoisovalerate (KIV), and α-keto-β-methylvalerate (KMV). The three substances are collectively known as branched alpha-ketoacids (BCKAs). Further, BCKAs produce acetyl-CoA through an irreversible rate-limiting reaction catalyzed by branched alpha-ketoate dehydrogenase (BCKDH) and subsequent reactions. Acetyl-CoA may be involved in the TCA cycle or other amino acid synthesis. Glutamine can be transported by SLC1A5 (ASCT2), LAT1 (L-type amino acid transporter), and xCT (SLC7A11). Glutamine is involved in glutathione (GSH) synthesis and cell REDOX homeostasis regulation in cytoplasm. In the mitochondria, glutamine produces Glutamate through a reaction catalyzed by glutaminase (GLS), which participates in the TCA cycle by producing α-KG by aminotransferase (ATs) and Glutamate dehydrogenase (GLUD).
l-Phenylalanine/l-tyrosine: The tetrahydrobiopterin-dependent hydroxylation of phenylalanine generates tyrosine. Catabolism of tyrosine generates the glucogenic Krebs-cycle intermediate fumarate and the ketogenic metabolite acetoacetate in five reaction steps. Complete oxidation of phenylalanine requires biopterin, ascorbate, thiamin, riboflavin, niacin, vitamin B6, pantothenate, lipoate, ubiquinone, iron, and magnesium. Tyrosine is also a precursor of catecholamines and melanin.
l-Tryptophan: Almost all tryptophan is eventually metabolized through a long sequence of reactions to alanine and two acetyl-CoA molecules. Alternative tryptophan derivatives of biological importance include serotonin, melatonin, nicotinamide adenine dinucleotide (NAD), and nicotinamide adenine dinucleotide phosphate (NADP). Adequate supplies of thiamin, riboflavin, vitamin B6, niacin, pantothenate, lipoate, ubiquinone, iron, and magnesium are necessary for normal utilization.
l-Methionine/l-cysteine: Methionine is converted into cysteine via homocysteine. The intermediate homocysteine is extensively remethylated to methionine in a folate- and vitamin B12–requiring reaction. Most cysteine is metabolized to pyruvate through several alternative pathways. Smaller amounts are converted into alanine and taurine. These metabolic pathways require thiamin, riboflavin, niacin, vitamin B6, pantothenate, lipoate, ubiquinone, iron, and magnesium. Disposal of the toxic sulfite requires molybdenum
l-Serine: Breakdown of serine can proceed with pyruvate as an intermediate, but most is used for the synthesis of glycine, cysteine, alanine, selenocysteine, and choline (via phosphatidylserine). These are then eventually catabolized. Complete oxidation requires thiamin, riboflavin, vitamin B6, niacin, lipoate, ubiquinone, iron, and magnesium.
l-Threonine: The mitochondria catabolize threonine to acetate and glycine. An alternative mitochondrial pathway can generate a spectrum of metabolites including acetol, lactaldehyde or d-lactate. Cytosolic metabolism leads to the glucogenic Krebs-cycle intermediate succinyl-CoA. Threonine breakdown uses thiamin, riboflavin, niacin, vitamin B6, folate, vitamin B12, pantothenate, biotin, lipoate, ubiquinone, zinc, iron, and magnesium.
l-Asparagine/l-aspartate: Asparagine can be deaminated to aspartate and then this amino acid is converted to the glucogenic Krebs-cycle intermediate oxaloacetate. Aspartate is a precursor of purine and pyrimidine nucleotide synthesis. Complete oxidation of either one depends on thiamin, riboflavin, niacin, vitamin B6, lipoate, ubiquinone, iron, and magnesium.
l-Glutamine/l-glutamate: Glutamine can be deaminated to glutamate, and then to the glucogenic Krebs-cycle intermediate alpha-ketoglutarate. Glutamine is a precursor of purine and pyrimidine nucleotide synthesis. Complete oxidation of either glutamine or glutamate depends on thiamin, riboflavin, niacin, vitamin B6, ubiquinone, iron, and magnesium.
l-Histidine: Most histidine is metabolized to glutamate. A minor alternative pathway generates imidazole pyruvate, imidazole acetate, and imidazole lactate, which are excreted. The methylhistidine from modified proteins and anserine cannot be utilized as an energy fuel. Histidine breakdown requires thiamin, riboflavin, vitamin B6, niacin, pantothenate, lipoate, ubiquinone, iron, and magnesium
l-Lysine: This cationic amino acid is broken down into two molecules of acetyl-CoA in 10–13 steps, depending on the pathway. Complete oxidation depends on thiamin, riboflavin, vitamin B6, niacin, pantothenate, lipoate, ubiquinone, iron, and magnesium.
l-Arginine: Breakdown of arginine proceeds via glutamate to alpha-ketoglutarate, which is metabolized further through the Krebs cycle or used for glucose synthesis. Arginine is also a direct precursor of creatine and nitric oxide. Thiamin, riboflavin, vitamin B6, niacin, pantothenate, lipoate, ubiquinone, and magnesium are needed for the utilization of arginine
Urea synthesis: Most of the nitrogen ingested with protein eventually ends up as urea in urine. Urea synthesis occurs mainly in the liver. Carbamoyl phosphate synthase I (EC6.3.4.16) in the mitochondrial matrix condenses ammonia and bicarbonate in an ATP-driven reaction. Mitochondrial ornithine carbamoyltransferase (OTC EC2.1.3.3) can then join l-ornithine and carbamoyl phosphate to form citrulline. The ornithine for this reaction comes from the cytosol, from where the mitochondrial ornithine transporter 1 (SLC25A15) shuttles it in exchange for citrulline across the inner mitochondrial membrane (Camacho et al., 1999). In the cytosol, citrulline is condensed with l-aspartate in an ATP-driven reaction (argininosuccinate synthase, EC6.3.4.5). Argininosuccinate lyase (EC4.3.2.1) then cleaves this intermediate into fumarate and l-arginine. The cycle is finally completed when the manganese-requiring enzyme arginase (EC3.5.3.1) cleaves l-arginine into urea and l-ornithine. While ornithine is mostly recycled during urea synthesis, it comes ultimately from de novo synthesis in the small intestine. Smaller amounts may be taken up from food sources
Ornithine synthesis from l-glutamate occurs in the kidneys, intestine, brain, and other tissues. Glycine amidinotransferase (EC2.1.4.1) in the kidneys, which is the rate-limiting enzyme for creatine synthesis, produces l-ornithine and guanidinium acetate from l-arginine and glycine. An alternative synthesis pathway, mainly in the small intestine, starts with the phosphorylation by mitochondrial gamma-glutamate 5-kinase (EC2.7.2.11). In humans, the same protein also catalyzes the subsequent reduction to glutamate gamma semialdehyde (glutamate gamma-semialdehyde dehydrogenase, EC1.2.1.41). Ornithine-delta-aminotransferase (EC2.6.1.13) can then complete ornithine synthesis by transferring the amino group from l-glutamate or a number of other amino acids to glutamate gamma-semialdehyde.
A constant proportion of l-glutamate in liver mitochondria is acetylated by the amino acid N-acetyltransferase (EC2.3.1.1). Since N-acetyl glutamate activates carbamoyl phosphate synthase, the concentration of l-glutamate sets the pace of urea synthesis.